Dong-fang Li1,
Rui-xiao Wang2,
Yong-xia Yan1,
Guo-liang Jin1,
Guo-hui Song2,
Deng-bin Ma3,
Li Guan2 ![]()
For correspondence:- Li Guan Email: guanligl@hotmail.com
Received: 5 July 2016 Accepted: 7 January 2017 Published: 25 February 2017
Citation: Li D, Wang R, Yan Y, Jin G, Song G, Ma D, et al. In-silico investigations into natural products as non-nucleoside DNA methyltransferase 1 inhibitors for treating epi-mutation in gastric cancer. Trop J Pharm Res 2017; 16(2):445-451 doi: 10.4314/tjpr.v16i2.25
© 2017 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To explore in silico methods to search for the best reported non-nucleoside DNA methyltransferase 1 (DNMT1) inhibitor of epimutation in gastric cancer.
Methods: A dataset of reported non-nucleoside DNMT1 inhibitors was used to target the active site of crystallized DNMT1 protein. Molecular docking simulations were carried out using AutoDock 4.2.6 l. The results were analyzed using Discovery studio visualizer.
Results: In silico analysis of known natural non-nucleoside DNMT1 inhibitors gave genistein as the top ranked compound with ΔG of -6.39 Kcal/mol. Further, the results indicated that epigallocatechin gallate and curcumin are poor non-nucleoside DNMT1 inhibitors, as the in silico data suggest that they failed to bind to the catalytic site of DNMT1.
Conclusion: The results indicate that genistein is the top rated compound for DNMT1 inhibition. Previous in vitro and in vivo work by other researchers seem to validate the findings of the study
Introduction
The inhibition of DNA methyltransferase1 (DNMT1) can be achieved by two inhibitory mechanisms: enzyme trapping achieved by nucleoside inhibitors and enzyme blocking by non-nucleoside inhibitors. A simple derivative of the nucleoside cytidine 5-azacytidine, is an archetypal DNMT1 inhibitor. The compound was first described more than four decades ago [1]. However, its demethylating activity was discovered later, in 1980 [2]. 5-azacytosine was shown to influence cellular differentiation by incorporation into DNA, and to acts as a nucleoside inhibitor [3]. However, this inhibitor was found to affect a variety of RNA functions and cause certain toxic effects [4]. Another DNMT1 inhibitor, 5-aza-2 ′ -deoxycytidine (i.e., decitabine), is a deoxyribose analogue of 5-azacytidine and has no effect on RNA function, but shows substantial toxic effects [5]. Another addition to the group of nucleoside DNMT1 inhibitors is zebularine [6], which is also a derivative of 5-azacytidine. This analog is more stable than others and its toxicity is less severe than the others [7].
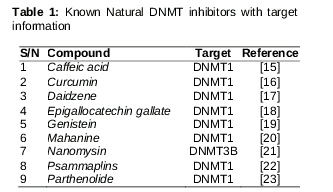
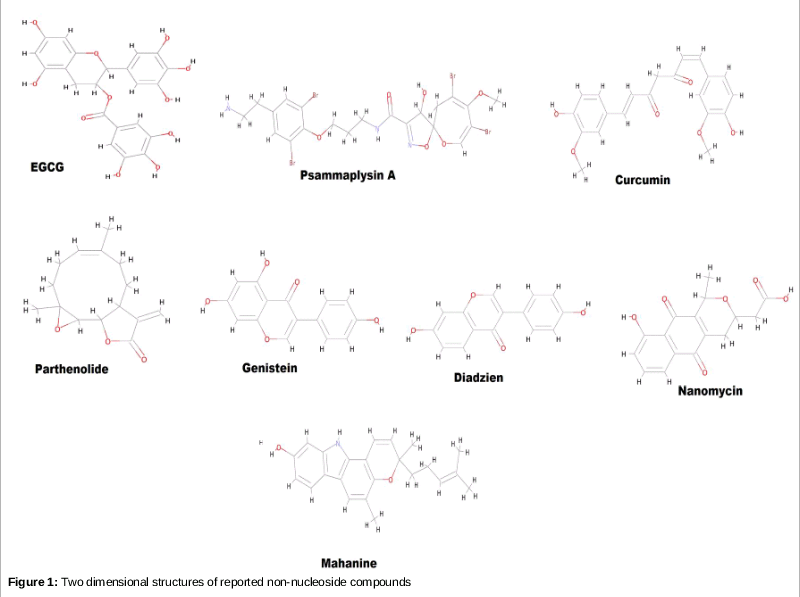
In all of the above nucleoside DNMT1 inhibitors, toxicity is the only factor that poses a substantial drawback for their development as viable therapeutic agents. To overcome this limiting factor, non-nucleoside DNMT1 inhibitors can be used as epi-drugs. A number of non-nucleoside compounds have been shown to inhibit DNMT1 activity, including Caffeic acid, Curcumin, Daidzene, Epigallocatechin gallate, Genistein, Mahanine, Nanomycin, Parthenolide, and Psammaplin. The third type of DNMT1 inhibitorconsists of oligonucleotides (hairpin loops and specific antisense oligonucleotides) such as MG98. Hairpin loops have been used as competitor substrates for DNMT1 in mouse erythroleukemia cells, and they have shown the ability to induce the expression of p16 TSG in human HT29 colan carcinoma cells [8]. Transfection of human HCT116 and SW48 colon cancer cell lines with antisense oligonucleotides against DNMT1 resulted in the demethylation and reactivation of expression in p16 gene expression [9].
Epimutation involving downregulation of important tumor suppressor genes is a common anomaly in gastric cancers [10]. Epi-drugs of nucleoside origin are in use for treatment; however, non-nucleoside epi-drugs are in developmental stage. In silico studies have been used for epi-drug discovery [11-13]; in the present study, we used a similar approach to examine the possible binding mode of reported non-nucleoside DNMT1 inhibitors using AutoDock.
Methods
Dataset preparation
Nine natural products, i.e, Caffeic acid, Curcumin, Daidzene, Epigallocatechin gallate, Genistein, Mahanine, Nanomycin, Parthenolide and Psammaplin, were selected for this study. The structures of these ten compounds are available from NCBI and were used for docking analyses. All were docked onto the active site, i.e., CYS1226, of DNMT1 using Auto Dock 4.2, and the result were analyzed using Accelrys discovery studio. The crystal structure of the catalytic site of human DNMT1 (accession: 3SWR) was retrieved from the protein databank (PDB). The retrieved structure was energy minimized using SPDB viewer at the default cutoff root mean square deviation (rmsd) of 0.30 Å using OPLS 2001 force field.
Molecular docking simulation
The selected compounds were analyzed using AutoDock to confirm the binding mode with the active site of the protein DNMT1 [14]. AutoDock energy values were calculated by characterization of intermolecular energy (which consists of van der Waals energy, hydrogen bonding energy, desolvation energy, and electrostatic energy), internal energy of the ligand, and torsional free energy. The docking energy was obtained from van der Waals energy and hydrogen bonding energy together, and the binding energy was built up from van der Waals energy and desolvation energy. Lamarckian genetic algorithm (GA) was considered for the run. For each ligand, 10 GA runs were set, with 27,000 maximum generations, gene mutation rate of 0.02, and crossover rate of 0.8. A grid of 60×60×60 points in x, y, and z directions was built centered around the catalytic site of CYS1226. All the visualizations were carried out using Accelrys Discovery studio visualization.
Results
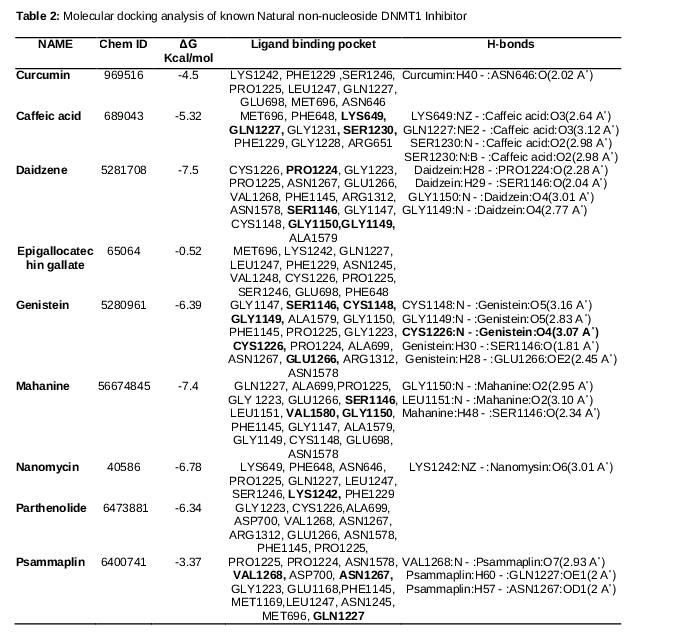
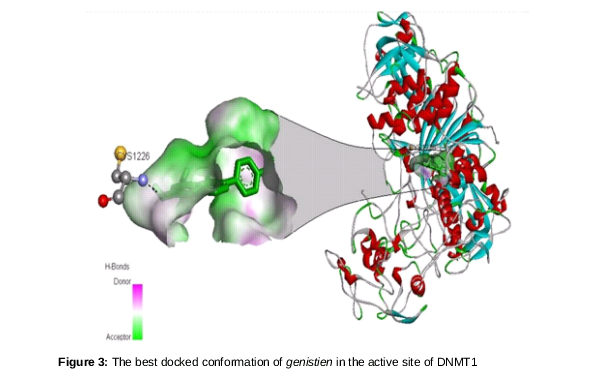
The top nine non-nucleoside compounds selected for this study are shown in , and their structures are shown in . The result generated by the molecular docking analysis of nine known natural compounds are shown in .
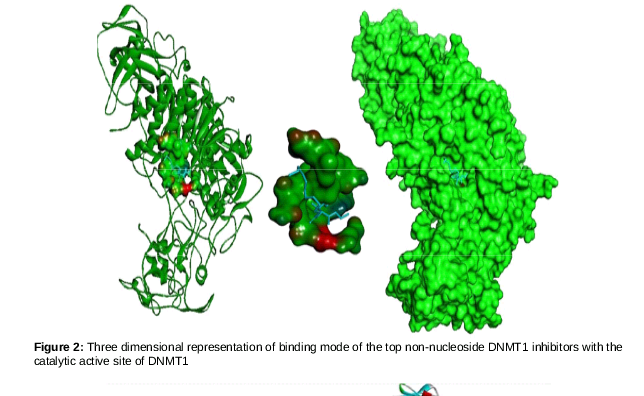
The data generated by analysis of these compounds with recently developed crystallographic structure showed that daidzene was the only compound interacting with the active site CYS1226, epigallocatechin gallate showed no interaction with the DNMT1 protein. The 3D plot of the interactions between the daidzen and the DNMT1 is given in .
Among the selected known non-nucleoside DNMT1 inhibitors, geneistein was the only compound that showed non covalent (hydrogen bond) interaction with the active site CYS1226 of the enzyme DNMT1. All other compounds except Epigallocatechin gallate showed interaction with other amino acids in the cytosine binding groove.
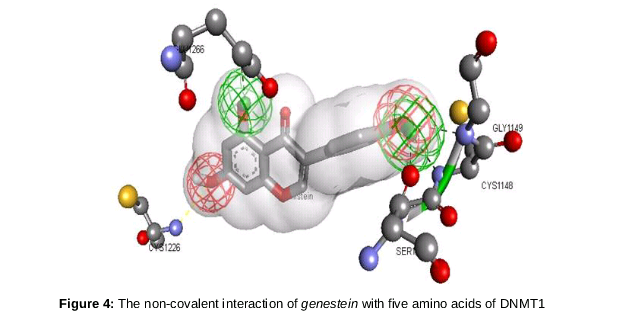
Genestein (chem. ID 5280961) has a ΔG of -6.39 Kcal/mol and forms five hydrogen bond with cytosine binding pocket of DNMT1. The binding pocket in this case comprises the following amino acids: GLY1147, SER1146, CYS1148, GLY1149, ALA1579, GLY1150, PHE1145, PRO1225, GLY1223, CYS1226, PRO1224, ALA699, ASN1267, GLU1266, ARG1312 and ASN1578.
The O4 position of the genestein interacts with CYS1226, and the bond formed between them has a distance of 3.07 A˚. spotlights on the binding scaffold of cysteine in DNMT1 and shows its interaction with genestein.
shows five different interactions. The first interaction is discussed is that of active site CYS1226 and Oxygen at 4th position of genestein. The oxygen atoms are represented by red mesh and interact at two positions- one has already discussed, and the other is the interaction of oxygen at the 5th position with GLY1149. In addition, three more interactions were observed, all of which, except that with GLU1266 arose from the same oxygen at position 5. The shortest interaction is that of 30th hydrogen of genistein and the oxygen atom of SER1146. This result generated by molecular docking of genestien onto the catalytic site of DNMT1 implies that this particular non-nucleoside DNMT1 inhibitor has a better chance of reverting epimutation than any other known natural non-nucleoside inhibitors. Further analysis indicated that Epigallocatechin gallate is over hyped non-nucleoside DNMT1 inhibitor, as the results showed that it fails to interact with the catalytic scaffold of DNMT1.
Discussion
Genistein is known to protect against colon cancer by increasing gene expression of several WNT signaling components by demethylating their promoter [24]. Genistein is also known to show effects similar to those of 5Aza-cytidine, which is currently undergoing phase 2 clinical trials as a treatment for prostate cancer. The demethylating activity of the green tea polyphenol Epigallocatechin gallate is controversial.
The experimental evidence supporting DNA demethylation in human cancer cells was limited to only indirect PCR analyses of only few cancer-associated genes [18] and the effect could not be reproduced by direct experimental approaches. In addition, it has been noted that Epigallocatechin gallate is a highly reactive compound that may inhibit DNA methylation non-specifically and/or indirectly. Further, Medina-Franco [11] suggested that Epigallocatechin gallate should not be considered as DNMT1 inhibitor.
Curcumin is other natural compound, the potential of which is over estimated [16]; subsequent studies on this particular compound also showed that it fails to induce promoter hypomethylation [11]. These previous result were validated by our molecular docking analysis of the crystallographic structure of the catalytic site of DNMT1 indicating that curcumin showed only one interaction i.e. with ASN649.All the compounds examined other that genestien showed weak interaction with the catalytic binding pocket of DNMT1, and the results of in vitro and in vivo studies also tend to agree with our in-silico results.
Conclusion
The findings of this study provide and atomic insight into the binding mode of known natural non-nucleoside DNMT1 inhibitors. The results generated are useful for identifying the top-ranking non-nucleoside DNMT1 inhibitor using in silico methodologies. Genistein shows the greatest inhibitory effect, and in vitro and in vivo studies of the same compounds by other researchers seem to validate these findings. Further, the results also indicate that epigallocatechin gallate and curcumin have been overestimated as potential non-nucleoside DNMT1 inhibitors as their in silico data generated by our approach suggest that they fail to bind to the catalytic site of DNMT1. Thus, this study provides a suitable, efficient and economically viable method to screen for novel non-nucleoside DNMT1 inhibitors.
References
Archives


News Updates